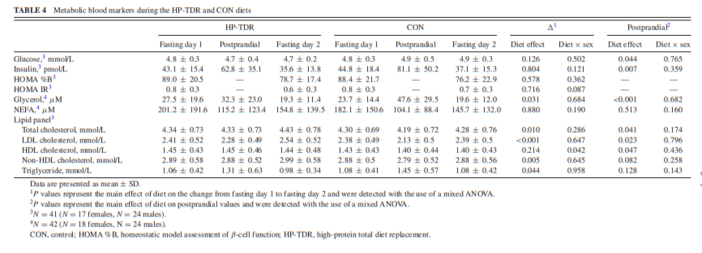
Metabolic blood markers
Metabolic blood markers assessed in a fasting state on days 1 and 2, and after lunch during the HP-TDR and CON diets are shown in Table 4. Glycerol (–4.2 ± 12.4 μM, P = 0.031) and triglyceride (–0.07 ± 0.23 mmol/L, P = 0.044) decreased more from fasting day 1 to fasting day 2 in the HP-TDR compared with the CON diet, and total, LDL, and non-HDL cholesterol blood concentrations increased more (0.10 ± 0.26 mmol/L, P = 0.010; 0.12 ± 0.18 mmol/L, P 0.05, respectively). On the other hand, this change was not different between the dietary interventions for glucose, insulin, HOMA %B, HOMA IR, NEFA, and HDL cholesterol, P >0.05.
There was a statistically significant interaction between diet and sex on the change in HDL cholesterol concentration (P = 0.042). In the HP-TDR diet, the HDL cholesterol concentration was greater in females compared with males (0.08 ± 0.03 mmol/L, P = 0.007). Moreover, the change in HDL cholesterol from fasting day 1 to fasting day 2 was significantly different between interventions in females (HP-TDR: 0.03 ± 0.03 mmol/L; CON: –0.01 ± 0.02 mmol/L; P = 0.043), but not in males (HP-TDR: –0.04 ± 0.02 mmol/L; CON: –0.05 ± 0.12 mmol/L; P = 0.525). There was no difference between sexes in the CON diet (P = 0.430).
Postprandially, glucose (–0.2 ± 0.5 mmol/L, P = 0.044), insulin (–19.1 ± 44.6 pmol/L, P = 0.007), and glycerol (– 16.8 ± 25.9 μM, P <0.001) blood concentrations were lower in the HP-TDR diet compared with the CON, and total, LDL, and HDL cholesterol concentrations were higher (0.12 ± 0.42 mmol/L, P = 0.041; 0.12 ± 0.34 mmol/L, P = 0.023; 0.06 ± 0.20 mmol/L, P = 0.047, respectively). There was no diet × sex interaction in any of the variables analyzed postprandially (all P >0.05).
The order in which participants received the dietary interventions did not affect any of the metabolic blood markers analyzed, P >0.05. A diet × sex × time interaction was also explored, and no statistically significant 3-way interaction was observed in any of the variables analyzed (Supplementary Table 2).
Discussion
The present inpatient metabolic balance study compared the effects of an isocaloric HP-TDR versus a CON diet on EE, macronutrient oxidation rates and balances, and metabolic blood markers in female and male healthy adults. The primary findings of this study were that compared with a standard North American dietary pattern, a HP-TDR led to higher total EE, increased fat oxidation, and negative fat balance (likely implying body fat loss) (25). The only diet × sex interaction observed was on HDL cholesterol concentration in the HP-TDR diet. These results highlight the impact HP-TDR consumption has on energy metabolism and metabolic blood markers of healthy adults and provides further insight into the potential role of this dietary strategy for weight management.
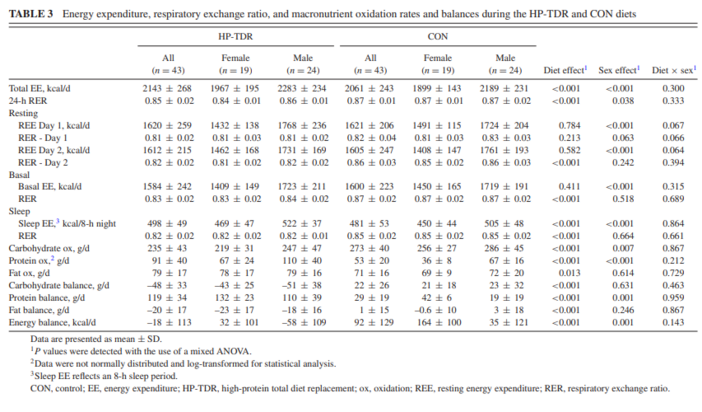
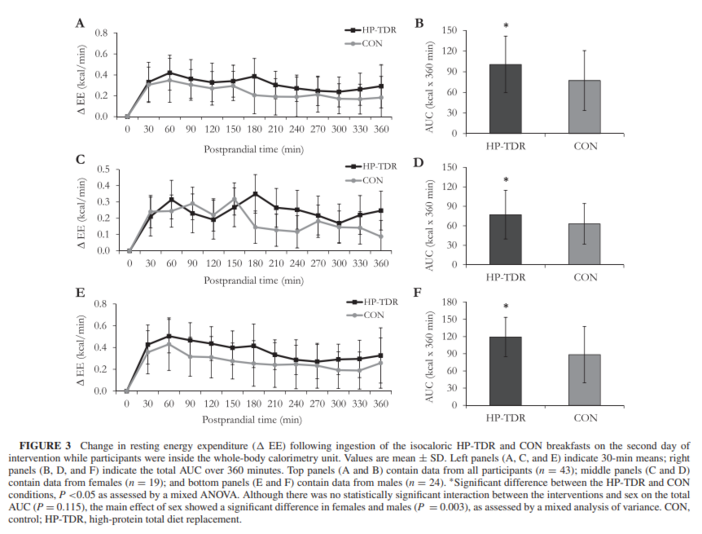
Regarding the components of participant’s EE, this study showed that consumption of the HP-TDR led to higher daily, sleep, and postprandial EE. Collectively, these results add to the discussion that a calorie is not just a calorie (26) and that isocaloric diets with a different proportion of macronutrients might offer a metabolic advantage (27–29), specifically an increase in EE and fat oxidation. There seems to be a consensus that the protein content of the diet can directly affect EE and substrate use (11, 30); however, the same is not true when it comes to the carbohydrate and fat contents (31, 32, 33). It is possible that energetic costs involved in the thermic effect of protein and the possible increase in protein turnover contributed to the observed increase in EE in this group (11, 30), which is concordant with the literature (13). On the other hand, 24 h after the start of the interventions, participant’s REE and basal EE did not differ between diets, contrasting previous findings (13). Interestingly, it seems that eucaloric HP diets are not able to change REE as it does with other components of EE, which can only be captured with the sophisticated measurement of energy metabolism (i.e., using a WBCU). Previous studies showing an increase or decline in REE with HP diets were long-term interventions in which participants were in negative or positive energy balance. A meta-analysis of randomized controlled trials revealed that HP diets reduced the decline in REE during weight loss, which has been potentially attributed to a retention of lean mass, although this has not been determined (34). In addition to that, overfeeding a HP diet for 8 wk has been shown to increase REE (227 kcal/d) and this result was associated with an accretion of 3.18 kg of lean mass (14). Due to our experimental design, lean mass and therefore REE were not expected to change, which is in line with current literature.
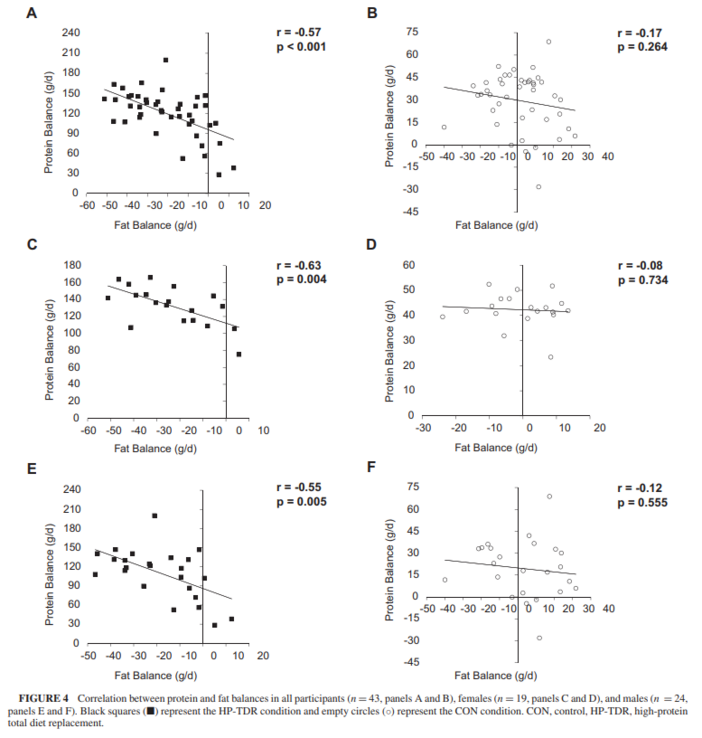
Over the years, experiments have shown that total body carbohydrate and protein content are tightly regulated by adjusting oxidation rates to intake levels, meaning that manipulating the intake of these macronutrients affects their oxidation rates to the same direction and extent (25, 35, 36). In this study, a HP-TDR led to a decrease in carbohydrate oxidation rate and an increase in protein oxidation rate, which is in line with this rationale since the HP-TDR intervention has a low-carbohydrate, HP content. Conversely, this autoregulatory process is nonexistent for fat oxidation, which seems to be mostly driven by the presence or absence of other macronutrients, markedly carbohydrate (25). The dynamic interactions between carbohydrate and fat oxidation started to be described almost 60 y ago (37) and have been continuously explored as more research is made available (38). As comprehensively discussed by Hue and Taegtmeyer (38) and illustrated by Prentice (25), the low-carbohydrate characteristic of the HP-TDR seems to be responsible for the increased fat oxidation observed with this dietary intervention. This result is further demonstrated by the lower 24-h RER observed in the HP-TDR diet. As a consequence of intake and oxidation rates in this study, participants consuming the HP-TDR experienced a decrease in energy and fat balances, likely implying body fat loss (25). In a classical inpatient experiment, Abbott et al. (36) demonstrated that in conditions of energy imbalance, fat stores are mobilized to balance the body’s energy budget, which is in agreement with results presented herein.
In this study, total, LDL, and non-HDL cholesterol blood concentrations increased more from fasting day 1 to fasting day 2 in the HP-TDR compared with the CON diet. Although change in these markers was statistically significant, the absolute values remained within the reference ranges for this population group. Jones et al. (39) demonstrated that the ingestion of dietary cholesterol causes feedback inhibition of cholesterol biosynthesis in humans. Considering that the content of dietary cholesterol of the HP-TDR intervention was almost 3 times lower than the content of the CON diet, it might be possible that the participants’ biosynthesis was upregulated in the HP-TDR, causing an increase in blood lipid concentrations. On the contrary, blood triglyceride concentration was lower in the HP-TDR diet compared with the CON diet. This fact can be mainly attributed to the low carbohydrate content of this dietary intervention (40), also supported by previous studies (41). Additionally, blood glycerol decreased more from fasting day 1 to fasting day 2 in the HP-TDR compared with the CON diet. Circulating glycerol has been shown to result mainly from hydrolysis of triglyceride stored in adipose tissue, and constitutes a major substrate for glucose homeostasis (42). The increased fat oxidation and negative fat balance observed in the HP-TDR group are both indicative of increased hydrolysis of triglycerides in adipose tissue, which might have greatly contributed to the use of this substrate as an energy source, reducing its circulating concentrations. A significant interaction between diet and sex on the change in HDL cholesterol concentration was found in the HP-TDR diet, in which females presented greater values compared with males. An analysis of 1.3 million patients revealed that the HDL cholesterol concentration is higher in females than males (43). This effect seems to be related to females’ increased endogenous (44) and exogenous estrogens (e.g., estrogen-containing contraceptives) (45). Considering that the HP-TDR contained soy isoflavones (46), which are natural estrogen-like compounds, it might be possible that it could have elevated the females’ estrogen concentrations, which accentuated the difference in HDL cholesterol concentration between sexes in the HP-TDR diet.
To date, studies investigating the effects of TDRs have been conducted in individuals with obesity and/or comorbidities in a state of negative energy balance with the main objective of weight loss (2, 4–9). The presence of several confounding variables in these studies, such as weight loss and comorbidities, hinders our understanding of the real physiological impact of TDRs. To our knowledge, this is the first study to compare a HP-TDR with a North American diet in healthy young adults of both sexes. In addition to being the first on the topic, this study has several strengths, including its crossover and rigorously controlled feeding design, allowing the detection of small diet effects on energy metabolism variables and metabolic blood markers. Moreover, the use of state-of-the-art technology, such as the WBCU, provided highly accurate and precise results, reflecting the real effects of the dietary interventions. In addition to the design and technology used, the study of both females and males allowed us to explore how different sexes respond to these dietary interventions.
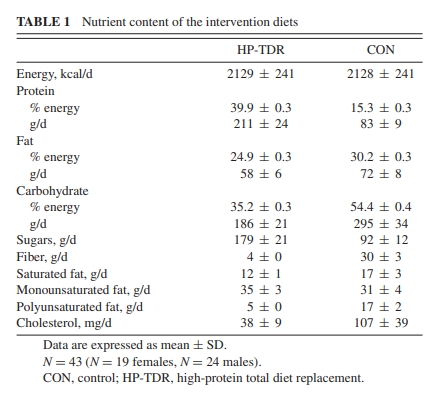
In this study, participants received isocaloric diets that were designed to mimic the North American dietary pattern and a nutritional product commercially available in many countries. For this reason, 1 or more macronutrients could not be kept constant in 1 dietary intervention while others were manipulated in the other intervention. When comparing the macronutrient distribution of the HP-TDR with the Acceptable Macronutrient Distribution Range (10), this dietary strategy can be characterized as HP and low-carbohydrate. Therefore, it is not possible to attribute any of the results observed in this study to a single macronutrient. In addition, this study has other limitations, including the specificity of the population being studied (i.e., healthy, young adults with a normal body weight) and the short-term intervention. These limitations restrict our ability to translate these results to other population groups and longer intervention periods. Therefore, future studies are needed to better understand the long-term effects of this dietary intervention on the physiology of healthy and diseased population groups.